
Palaeoenvironmental conditions can be reconstructed from microfossils preserved in sediments using the relationship between species and the environment. Usually the species’ relationships with the environment — their niches — are insufficiently constrained by experimental data, so we are forced to rely upon observations of species abundances and environmental conditions in a modern calibration set and assume that we can make useful inferences of the niche from these data.
Sometimes this assumption fails. This post tells of one such case.
I did my PhD, and some subsequent work, trying to infer climate from diatom assemblages in sediment cores from closed-basin lakes in Ethiopia, and later Tanzania. Closed-basin lakes are lakes that don’t have a river flowing out of them (or groundwater leakage), and should be sensitive to climate change: deep and relatively fresh when the climate is humid; and shallow and saline during arid periods. Diatoms, algae with beautiful siliceous cell walls or frustules that preserve well in most lake sediments, are sensitive to salinity and water depth and so should be possible to use to constrain past changes in hydrology. Françoise Gasse and coworkers (Gasse et al 1995) developed a transfer function to reconstruct conductivity (a variable related to salinity) from diatom assemblages. I used this transfer function in my thesis and subsequent work.
The first paper I wrote was on a 6500-year long diatom stratigraphy from Lake Awassa, a large but fairly shallow caldera lake in the Ethiopian rift valley. Even though the lake has no out-flowing rivers, it is fairly fresh, suggesting that there is groundwater leakage, probably to the north. I was expecting to find evidence that Lake Awassa was deeper and fresher during the early Holocene, which was more humid in most of North Africa, with lakes scattered across the Sahara. I was hoping to be able to make a continuous climate reconstruction across the end of the termination of the early Holocene humid period – was it an abrupt or gradual change? Instead, the diatom-conductivity model suggested that the lake was more saline during two phases of the mid-Holocene, before the expected termination of the humid period. Either the timing and nature of the termination inferred from several sites across North Africa were incorrect, or there was something funny going on in Lake Awassa. I decided that the latter was more plausible.
Both of the apparently saline phases in Lake Awassa occurred immediately after very fine-grained tephra layers, so I investigate the potential for a volcanic origin for the saline phases. I found sufficient evidence from other sites to make what I thought was a plausible case for pulses of saline groundwater somehow related to the volcanism making the lake more saline.
One of the main salinity indicators in my Lake Awassa core was Thalassiosira faurii. Most Thalassiosira are marine species, but some live in, often brackish, lakes.

Thalassiosira faurii (Roubeix et al 2014)
Gasse et al (1995) estimated the conductivity optimum of T. faurii to be ~9000 μScm-1, calculated by weighted-averaging. With the European Diatom Database (EDDI) we can look at the data behind this estimate, and plot them (you might need to enable java for unsigned applets – sorry I wrote/borrowed the java code for these plots long before security holes in java were known).
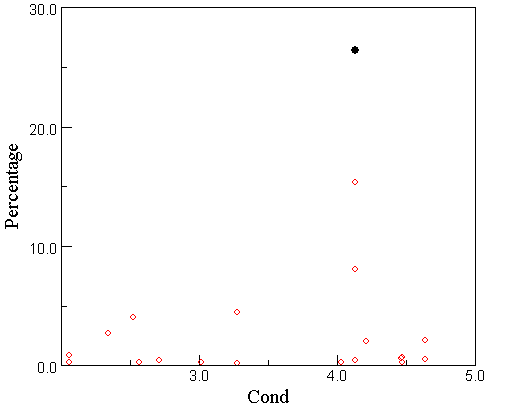
T. faurii relative abundance against salinity in the African diatom calibration set
The marked sample and the other two samples with a high relative abundance of T. faurii are all from a salt swamp in Niger. Without these samples, the optimum would be somewhat lower. This dependence on one site, where T. faurii does not reach the maximum relative abundance (60%) found in Late Awassa, is not ideal.
Roubeix et al (2014) investigated the salinity niche of T. faurii collected from Lake Langano, another rift-valley lake 60 km to the north of Awassa, by measuring its growth rate when grown at different salinities under controlled conditions in a laboratory.

Growth rates of Thalassiosira faurii (black diamonds) and Anomoeoneis sphaerophora (open diamonds) versus conductivity of the culture medium. The dashed line shows the variations of pH between cultures. Vertical bars represent the standard deviations of the replicates. A star means that the cells were still alive after 15 days, whereas a cross means that they were all dead. The two triangles indicate the conductivity and pH of lake water from which the diatoms were isolated.
At least the clone grown by Roubeix et al (2014) did not have an conductivity optimum of ~9000 μScm-1; it died at conductivities this high. The optimal conductivity was estimated at just 400 μScm-1. This is the optimum in the absence of any competition, the realised optimum in lakes many be somewhat higher, but is unlikely to be much above ~1000 μScm-1, as by ~2000 μScm-1 there is no growth. Roubeix et al use this new information to reinterpret the conditions in Lake Abiyata, a saline lake adjacent to Langano, during the Younger Dryas. Previous work by Chalié and Gasse (2002) had inferred saline conditions during this interval.
Roubeix et al also question my reconstruction from Lake Awassa. If T. faurii has a much lower conductivity optimum, then the conductivity spikes I reconstruct in the mid-Holocene may be an artefact. While I recognise that my proposed mechanism of pulses of saline hydrothermal water is speculative, I’m not entirely convinced that I was wrong as the salinity spike is not based on the presence of T. faurii alone. Instead, I found T. faurii in association with the saline indicators Navicula elkab (optimum ~14000 μScm-1) and T. rudolfii (optimum ~11400 μScm-1). Both these conductivity optima estimates seem to be well supported by data from a variety of sites.
The discrepancy between the old observational and new experimental optimum of T. faurii may reflect taxonomic problems. Perhaps T. faurii is not a single species but a suite of morphologically similar cryptic species with different salinity niches, with the diatoms from the salt swamp in Niger representing one end of the spectrum and the clone from Langano representing the other. Unless morphological characteristics that distinguish salt-tolerant from non-tolerant variants of T. faurii, it will not be possible to guarantee which variant is present in the palaeoecological records and any reconstruction will be rather uncertain.
Cryptic species are known in several of the taxonomic groups used for reconstructing climate, so there is a risk of cryptic species with different ecological preferences may be a widespread problem in palaeoecology.
Part of the problem relates to the lack of external data. If I examine a pollen calibration set and find the optimum temperature for lime (Tilia) to be 5°C, there is enough literature on the ecology of Tilia to recognise that there is a problem. With diatoms, dinoflagellate cysts and many other groups, there is little or no literature on the ecology of the different species, so we cannot easily recognise when the optima we calculate are spurious. Yet another reason why care is required when using transfer functions to reconstruct palaeoenvironmental conditions.
Gasse, F., Juggins, S. & Ben Khelifa, K. 1995. Diatom-based transfer functions for inferring past hydrochemical characteristics of African lakes. Palaeogeogr. Palaeoclim. Palaeoecol. 117: 31–54.
Roubeix, V., Chalié, F. & Gasse, F. (2014) The diatom Thalassiosira faurii (Gasse) Hasle in the Ziway–Shala lakes (Ethiopia) and implications for paleoclimatic reconstructions: Case study of the Glacial–Holocene transition in East Africa. Palaeogeogr. Palaeoclim. Palaeoecol. 402: 104–112.
 @richardjtelford
@richardjtelford
